超高速X線ムービー撮影により昆虫の羽ばたき機構を解明(プレスリリース)
- 公開日
- 2013年08月26日
- BL40XU(高フラックス)
2013年8月26日
公益財団法人 高輝度光科学研究センター
|
高輝度光科学研究センター(JASRI)は、羽ばたき中の昆虫の胸部に強力なX線を当て、毎秒5000コマという超高速X線ムービーを記録することで、昆虫の速い羽ばたきを可能にする筋肉の分子機構を明らかにすることに成功しました。 (論文) |
研究の背景
進化した昆虫(ハチ、ハエ、蚊、甲虫など)は非常に速く羽ばたくことができ、蚊の場合は1秒間に500回も羽ばたきます。脊椎動物の筋肉は1回の神経指令により1回収縮しますが、これと同じやり方で1秒間に500回も羽ばたくのは不可能です。そこで、進化した昆虫では飛翔筋(羽を動かす筋肉)が収縮したまま自励振動を行なうことで高い羽ばたき周波数を実現しています。この自励振動は、飛翔筋の「伸張による活性化」(筋肉を外から引っ張ると大きな力を出して引っ張り返す性質)という機能によって起こされます。図1にその仕組みを解説します。
進化した昆虫の飛翔筋は間接飛翔筋と呼ばれ、羽を直接動かすのではなく胸部の外骨格を変形させることで間接的に羽を動かします。胸部には2種の拮抗する飛翔筋があり、1つは前後方向に走り(DLM※2)、もう1つは背腹方向に走ります(DVM※2)。これらは片方が縮むともう片方が引き伸ばされる関係にあり、引き伸ばされた方は「伸張による活性化」により相手を引っ張り返しますから、お互いを引っ張り返すことで振動を持続することができるのです。これは筋肉自身のもっている性質を使っているので、振動ごとに神経が指令を送る必要はありません。
この「伸張による活性化」は他の筋肉にもみられる現象ですが、昆虫飛翔筋には特に顕著にみられます。その顕著さの故に古くから研究者の関心を集めてきましたが、
その分子機構は長年の研究にもかかわらず不明のままでした。最近は昆虫には特殊な飛翔筋特異的タンパク質(昆虫の体内にある多数の筋肉のうちで、飛翔筋にしか発現しない)が多く発現していることが明らかになり、これらの飛翔筋特異的タンパク質が「伸張による活性化」を起こすという説がかなり有力視されている状況でした。
研究内容と成果
本研究グループは今回、大型放射光施設SPring-8の中でも、他の標準的なビームラインより1000倍明るいBL40XUビームラインのX線を使い、羽ばたいているマルハナバチの胸部に当てて生じるX線回折像を世界最高速の毎秒5000コマの速さで記録することに成功しました。マルハナバチの羽ばたきは毎秒120回(羽ばたき1回が8ミリ秒)ですから、羽ばたき1回を40コマの高精度で記録でき、精密な解析が可能になりました。
これまでにアメリカの放射光施設APSで、羽ばたいているショウジョウバエの飛翔筋からX線回折像を記録した例はありました。しかしこのときは羽ばたき1回につき8コマの撮影速度で、2つの拮抗飛翔筋のうちDLMだけの記録でした。
今回の実験の特徴は、2つの拮抗飛翔筋が重なるところを狙ってX線を当てることで2つの拮抗飛翔筋からの回折像を同時記録したということです。これによって2つの拮抗飛翔筋がどういうタイミングで作動しているかが明らかになりました。2つの拮抗飛翔筋は実際には図1のように直交してはおらず、約60度の角度をなしていますが、動作は完全に逆位相(ほぼ正確に180度ずれている)ことが分かりました。
毎秒5000コマの回折像撮影には最新式の高速CMOSビデオカメラ※3(1024x1024ピクセル)を使いました。ポイントはこのビデオカメラを2台同時に使ったことです(図2)。1台はX線回折像撮影に使い、もう1台はマルハナバチの羽ばたきそのものを記録しました。マルハナバチを撮影したムービーの1コマを図3に示します。2台のカメラはマスタースレーブ方式※4で接続されているので完全な同期がとれ、回折像と羽の位置を1対1に対応させることができました。
以上の測定により記録されたX線ムービーの1コマが図4です(Scienceのサイトには論文のサプリメント※5として実際のムービーが公開されます)。これには2つの拮抗飛翔筋からの回折像が重なって記録されていますが、この図ではDLMの向きが縦になるように角度を回転させて表示しています。多数のスポット(反射)が見え、それぞれが筋肉内のタンパク分子の構造に関する情報を含んでいます。実際のムービーではこれらのスポットが明るくなったり暗くなったりしていますが、これは各タンパク分子の構造が変化していることを表します。また各スポットの位置を精密に測ることで、筋肉の長さや発生張力が羽ばたきにつれてどう変化しているかも知ることができます。例えば長さ変化はほぼ正確に正弦波振動でしたが、張力の変化はかなり歪んでいることが分かりました。
今回得られた情報でとりわけ重要なのが、図4の中で111と示されたスポットで、これが「伸張による活性化」の起こる直前に明るくなることが分かりました。すなわちこれが「伸張による活性化」の引き金になる構造変化を反映している可能性が高いのです。回折学的な検討の結果、この信号は、収縮タンパクのミオシンがもう一つの収縮タンパクであるアクチンに結合したままねじれるような変形をすると説明できることが分かりました(図5)。すなわち、アクチンに結合して力発生の準備段階にあるミオシンが伸張により変形を受けることで力発生を開始することが「伸張による活性化」の分子機構であると推定されます。
疲労して張力の低下した脊椎動物の骨格筋を引き伸ばすと非常に大きな力を出して抵抗しますが、この伸張による活性化も同様の分子機構で起こっているという論文を同じ著者のグループが1995年に出版しています。つまり昆虫飛翔筋と脊椎動物骨格筋は同様の分子機構によって引っ張りによる張力増強を行なっているということになります。昆虫飛翔筋は、飛翔筋特異的なタンパク質を発現するのではなく、脊椎動物にもある一般的な筋肉の性質を利用することによって「伸張による活性化」を実現したと考えられます。
今後の展開
最初に述べたように「伸張による活性化」は程度の差こそあれ種々の筋肉に広く見られる現象で、特に心筋ではその機能に対して重要な役割を担っていると考えられています。昆虫飛翔筋に顕著にみられる「伸張による活性化」が特殊な筋肉の特殊な性質ならば、その研究の意義も限られたものになるでしょうが、今回の研究で脊椎動物の筋肉との共通性が明らかになったことで、昆虫飛翔筋は脊椎動物の骨格筋や心筋の働きをよりよく理解するためのモデル材料として役立っていくことと思われます。
また応用的な面として、進化した昆虫飛翔筋は自励振動によって中枢を個々の振動を制御する負担から解放するというデザインで、これにより昆虫は全体を小型化して生息環境を広げることができたと考えられます。人間がロボット等の機器を開発する際にも、同様のデザインを用いて末端のアクチュエータの制御をアクチュエータ自身に行なわせることでCPUの負担を減らし、機器の小型化につなげることができると考えられます。
ここで紹介した研究は、日本学術振興会(No. 23612009)による科学研究費補助金の助成を受け、SPring-8の利用研究課題として行われました。
《参考図》
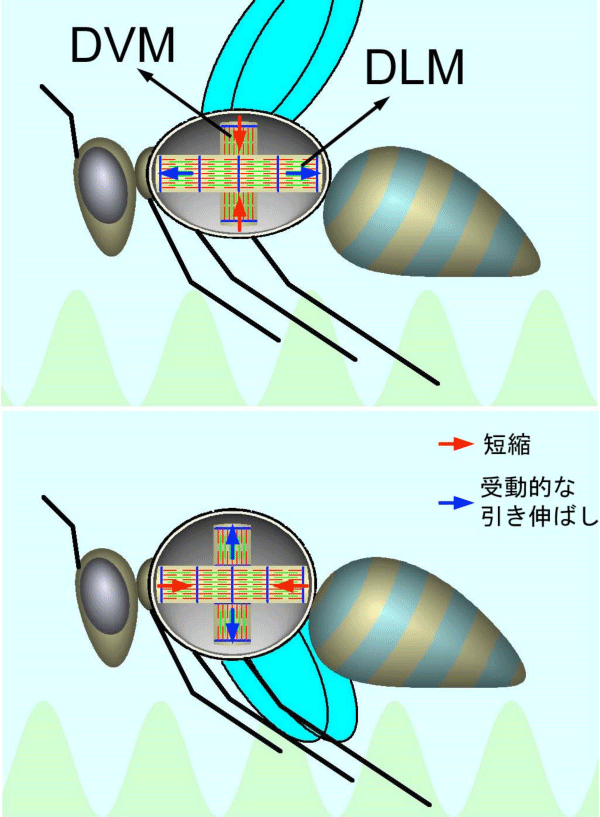
2種の拮抗する筋肉(DLM、DVM)があり、片方が縮むと、胸部外骨格の変形を介して他方が引き伸ばされる配置になっている。DLMとDVMは互いに引き伸ばしあうことから連続して収縮が起こり、羽ばたきを引き起こす。
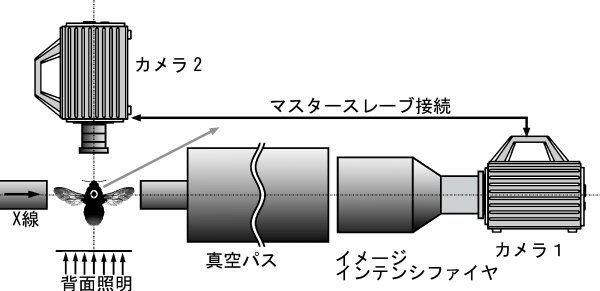
2台のカメラをマスタースレーブ接続し、X線回折像とハチの像を同時記録する。

ソフトウェアにより羽の位置を認識している(黄色の丸)。
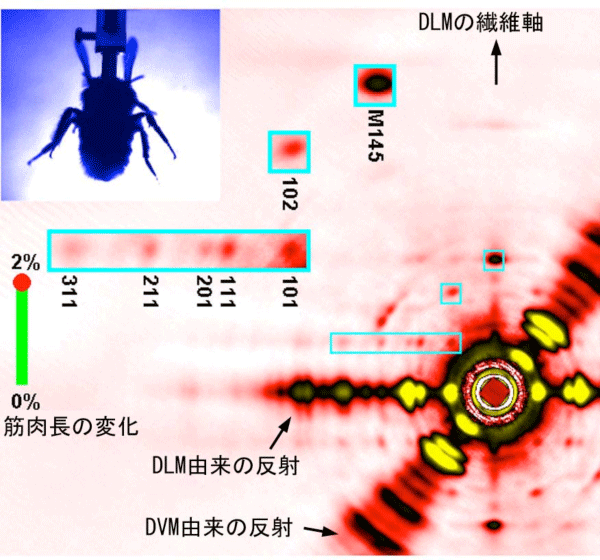
DLMの軸が垂直になるように回転させた上で、実験に用いた全てのハチのデータを加算したもの。青枠で囲った部分(全てDLM由来)は拡大して表示してある。
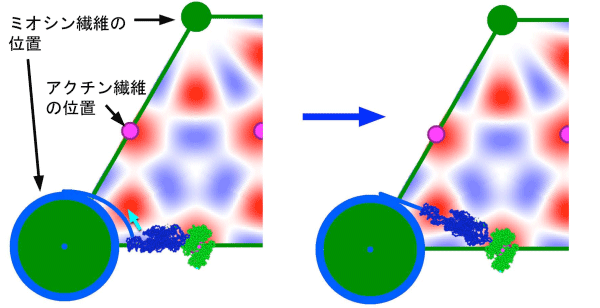
筋肉の横断面を拡大したもので、青色の不定形の物体がミオシン分子、緑の不定形の物体がアクチン分子を表す。ミオシンが左図から右図へとねじれるような動きをすると伸張による活性化に先立つ111スポットの強度変化を説明できる。
図2-5はScience誌より改変。
《用語解説》
※1 大型放射光施設SPring-8
兵庫県の播磨科学公園都市にある、世界最高の放射光を生み出す理化学研究所の施設、その運転管理はJASRIが行っています。SPring-8の名前はSuper Photon ring-8GeVに由来。放射光とは、電子を光とほぼ等しい速度まで加速し、電磁石によって進行方向を曲げた時に発生する、細く強力な電磁波のこと。SPring-8ではこの放射光を用いて、ナノテクノロジー、バイオテクノロジーや産業利用まで幅広い研究を行っています。
※2
DLM
英語のDorsal Longitudinal Muscleの略。
DVM
英語のDorso-Ventral Muscleの略。
Dorsal、Dorso-は背中側、Ventralは腹側、Muscleは筋肉の意味で、いずれも日本語訳はない。
※3 高速CMOSビデオカメラ
撮像素子としてCMOS(Complementary Metal-Oxide Semiconductor)を用いたビデオカメラ。従来のCCD素子(Charge-Coupled Device)を用いたものと比較して、各画素にアンプを設置できるため高速撮影が可能。
※4 マスタースレーブ方式
主人と奴隷の意味で、1台目の機器のクロックで2台目を制御する接続方式。
※5 サプリメント
論文のページ制限を越える内容や動画など雑誌に印刷できない内容を補助的なコンテントとして雑誌のウェブサイトに掲載したもの。
|
《問い合わせ先》
(SPring-8に関すること) |
- 現在の記事
- 超高速X線ムービー撮影により昆虫の羽ばたき機構を解明(プレスリリース)