スプリングエイトの高輝度X線が明らかにした昆虫飛翔筋の構造進化(プレスリリース)
- 公開日
- 2005年12月14日
- BL40XU(高フラックス)
平成17年12月14日
(財)高輝度光科学研究センター
|
財団法人高輝度光科学研究センター(吉良 爽理事長)の岩本裕之主幹研究員らの研究グループは、大型放射光施設スプリングエイトの高フラックスビームラインBL40XUの高輝度X線にマイクロビーム・極低温回折法などの先端測定技術を組み合わせ、昆虫の羽ばたきに用いられる飛翔筋が構造的に進化した過程を明らかにしました。 |
1.背景
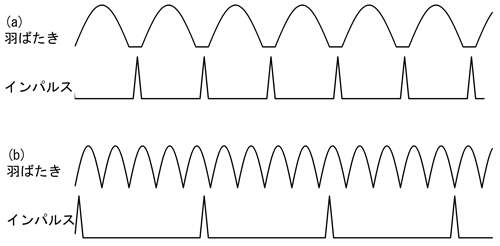
昆虫は地球上で最も種類数の多い動物群ですが、その繁栄の秘密は小型であることと飛翔能力にあるといっていいと考えられます。その飛翔を担う筋肉「飛翔筋」はヒトの骨格筋と同じ「横紋筋」(図1)に分類されますが、その動作方式には2種類あります。1つは「同期型」と呼ばれ、バッタのように比較的原始的な有翅昆虫に見られます。この方式では神経の興奮(インパルス)1回ごとに飛翔筋が収縮弛緩を行い、羽ばたきが1回起こります(図2a)。また大きく(10%以上)短縮することができます。これらの特徴はヒトの骨格筋と共通です。この方式で対応できる最大の羽ばたき頻度は毎秒100回程度と言われています。しかし小型の昆虫はこれ以上の羽ばたき頻度を必要としており、それは「同期型」とは異なる動作方式により実現しています。
もう1つの方式は「非同期型」といって、ハチやハエのような進化した昆虫に見られます。この方式では少ないインパルスで飛翔筋を収縮状態に保ち、胸部外骨格との共鳴によって飛翔筋を振動させて高い羽ばたき頻度を実現します。この場合、羽ばたきに比べてインパルスの頻度はずっと少ないです(図2b)。飛翔筋の振動に伴う長さ変化はわずか3%程度です。小さな長さ変化によって張力を正確にコントロールするには、筋節(1)中の収縮タンパクがよほど規則正しく並んでいないといけないと考えられます。実際、「非同期型」の飛翔筋の筋節を形作る収縮タンパク(アクチンやミオシンなど)の並び方は、ほとんど結晶と見なせるくらい非常に規則性の高いものであることが従来のX線回折実験や電子顕微鏡観察から分かっていました。これらの特徴はヒトの骨格筋にはないもので、「非同期型」の飛翔筋は飛ぶ目的のために特殊な進化を遂げたものであることがわかります。
2.今までの研究経緯
典型的な「非同期型」昆虫であるマルハナバチの1本の筋原繊維(2)に細いX線(マイクロビーム)を当てたところ、さらに驚くべきことがわかりました。筋節のタンパク結晶の格子面が筋節内だけでなく、筋原繊維の全長(約3mm、筋節1000個分)にわたって厳密に揃っていたのです(平成14年8月6日付のプレスリリース)。つまり、3mmもある筋原繊維の全体が1個の巨大タンパク単結晶とみなせるわけです。タンパクの原子構造を決めるために研究者はタンパク結晶を人工的に作りますが、これでも大きさは1mm以下です。昆虫はこれより大きな、しかも働く結晶を体の中に作ってしまうのです。
通常のX線回折実験ではX線を筋細胞の長軸に垂直に当てます。これに対して、上の実験では径2マイクロメートル程度に絞ったX線マイクロビームを筋細胞の長軸に平行にに当てました(エンドオン照射)。こうすることで、筋原繊維を取り出さずに、筋細胞の中に多数ある筋原繊維の1本だけにX線を当てて回折像を記録することができます。しかし当時は生の試料にそのままX線を当てましたので、照射ダメージが生じたり、長時間の記録の間に試料が乾いたりする問題がありました。そこで当研究グループは、試料を急速凍結し、極低温(−199℃)でX線マイクロビームによる生体試料の回折実験をおこなう方法を開発しました(本年6月29日付けのプレスリリース「単一筋原繊維中のただ1個の筋節からX線回折像記録に成功」参照)。これにより試料はX線照射ダメージに格段に強くなり、また測定中に試料が乾燥することもないので長時間安定に測定できるようになりました。
3.今回の研究成果
さて、先に述べましたようにマルハナバチ飛翔筋の筋原繊維は巨大単結晶型であることが分かったのですが、次の疑問は、この巨大単結晶型の筋原繊維が昆虫の進化の過程でどのように生じてきたかです。これを調べるため、原始的なものから進化したものまで、なるべく多くの種類の昆虫から飛翔筋を取り出し、筋原繊維のX線回折像を記録しました。方法は上に従って飛翔筋の筋原繊維を急速凍結し、X線マイクロビームをエンドオン照射するものです。
こうして各種昆虫の飛翔筋から記録した筋原繊維の回折像の例をいくつか示します(図3)。ハチ目(もく)、ハエ目、甲虫目、カメムシ類(カメムシ目の一亜目)の回折像はいずれもきれいな六角格子状になり、筋原繊維が巨大単結晶型であることがわかります。これらの昆虫は全て「非同期型」です。カミキリムシのような大型の甲虫は羽ばたき頻度も低く、「非同期型」動作は必要ないと思われますが、それでも回折像はきれいな六角格子状でした。最初に「非同期型」動作を獲得した祖先は小型の昆虫でしたが、その後大型種に進化しても飛翔筋の最初のデザインは忠実に受け継がれたといわれています。調べた限りでは全ての「非同期型」昆虫が巨大単結晶型の筋原繊維を持っていました。
それに対して、チョウ目やバッタ目など、多くの「同期型」の目では回折像がぼやけた同心円のように見え、格子面が全くそろっていないことが分かります。しかしカメムシ類と同じ目ながら「同期型」のセミ科では部分的な格子の規則性が見出され、まれに六角格子的な回折像がとれることがあります。この昆虫は進化の途上にあるか、あるいは大型化に伴って「非同期型」の特徴を失いつつあると考えられます。興味深いのは、有翅昆虫で最も原始的と考えられているトンボ目のイトトンボ類が六角格子的な回折像を示すことです。トンボ目の昆虫は、飛翔に強く依存した生活形態をとります。また同じく「同期型」のチョウ目の中でもスズメガ科だけは、多数のスポット状の反射からなる回折像を示します。これは解析すると、幾つかの六角格子の回折像が重なっていることが分かります。スズメガは、空中に静止しながら吸蜜するような高度の飛翔をします。
以上のように、巨大単結晶型の筋原繊維を持つものは概ね「非同期型」の昆虫に一致しますが、「同期型」の昆虫でも飛翔に強く依存した生活形態を持つものの一部はタンパク配列に部分的な規則性を示すことがわかりました。恐らく巨大単結晶型の筋原繊維は力の伝達特性がよく、進化上必要とされるときには独立に生じうるものであると考えられます。また、「非同期型」の昆虫でも足の筋肉の回折像は「同期型」の昆虫と同様なので、巨大単結晶型の構造が飛翔筋に特有のものであることがわかります。最後に、各種昆虫の飛翔筋動作タイプと筋原繊維の結晶性を表1にまとめます。
4.意義
今回の成果の意義は、スプリングエイトで生まれた測定技術「高輝度X線マイクロビームと急速凍結法の組み合わせ」が初めて生んだ科学の成果ということです。それは昆虫という、地球上で最も成功した動物群の進化の過程の一端を知り得たことです。体の小型化により、生物の機能の効率を極限まで高めたのが現在の昆虫の姿と思われます。その一例が「非同期型」飛翔筋の採用で、収縮弛緩に伴う無駄なエネルギーの使用を排しました。飛翔筋の構造もそのために最適化されましたが、その1つの答えが筋肉を巨大単結晶化することなのだと考えられます。昆虫が何億年の歳月をかけて獲得した効率化には、人間が小型デバイスを設計したりする上で参考にすべき点が多くあるのではないかと思います。本研究は文部科学省科学技術振興調整費及び科学研究費補助金No.15500294により支援を受けました。
<参考資料>
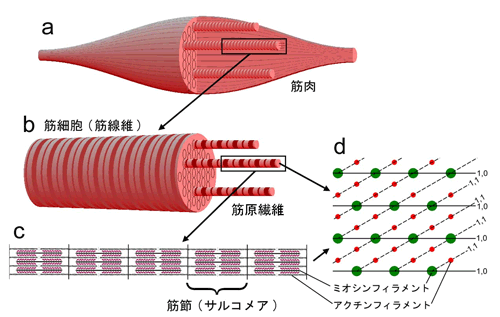
a:全筋。多数の筋細胞(筋線維)が集まってできている。
b:筋細胞。直径50 - 100マイクロメートル。直径1 - 2マイクロメートルの筋原繊維が多数集まってできている。昆虫の場合は筋細胞、筋原繊維とも太めである。
c:筋原繊維の拡大図。長さ2 - 2.5マイクロメートルの筋節(サルコメア)が多数直列に並んでいる。この中で収縮蛋白ミオシンとアクチンのフィラメントが六角格子状に配列している(d)。1個の筋節中では格子の向きは揃っている(単結晶)。d 中の数字(1,0,1,1)は特定の格子面を表わす指数。
a:同期型、b:非同期型

a:マルハナバチ(ハチ目)、b:フトハチモドキバエ(ハエ目)、c:ウリハムシ(甲虫目)、d:クロカミキリ(甲虫目)、e:アオクサカメムシ(カメムシ目)、f:ハルゼミ(カメムシ目)、g:エゾギクキンウワバ(チョウ目)、h:ホシヒメホウジャク(チョウ目スズメガ科)、i:ショウリョウバッタ(バッタ目)、j:ミヤマカワトンボ(トンボ目)
目 |
動作タイプ |
筋原繊維 |
ハチ目 |
非同期型 |
巨大単結晶型 |
ハエ目 |
非同期型 |
巨大単結晶型 |
甲虫目 |
非同期型 |
巨大単結晶型 |
カメムシ目カメムシ類 |
非同期型 |
巨大単結晶型 |
カメムシ目セミ科 |
同期型 |
中間型 |
アザミウマ目 |
非同期型 |
巨大単結晶型 |
チャタテムシ目 |
種による |
巨大単結晶型? |
チョウ目 |
同期型 |
結晶性低い |
チョウ目スズメガ科 |
同期型 |
中間型 |
シリアゲムシ目 |
同期型 |
結晶性低い |
トビケラ目 |
同期型 |
結晶性低い |
ヘビトンボ目 |
同期型 |
結晶性低い |
アミメカゲロウ目 |
同期型 |
結晶性低い |
ゴキブリ目 |
同期型 |
結晶性低い |
カマキリ目 |
同期型 |
結晶性低い |
カワゲラ目 |
同期型 |
結晶性低い |
カゲロウ目 |
同期型 |
結晶性低い |
トンボ目 |
同期型 |
中間型 |
ハチ目、ハエ目などの足 |
同期型 |
結晶性低い |
ウサギ骨格筋 |
同期型 |
結晶性低い |
表1 各昆虫飛翔筋の動作タイプと筋原繊維構造
<用語解説>
(1) 筋節:
サルコメアともいう。筋収縮の機能上の最小単位で、径、長さともに2-3マイクロメートル程度。この中でミオシンフィラメントとアクチンフィラメントが滑りあって力を発生する。
(2) 筋原繊維:
上記の筋節が直列に多数つながってできた径1-3マイクロメートルの繊維で、1本の筋細胞(筋線維、径100-300マイクロメートル)の中にはこれが多数含まれる。
|
(本研究に関する問い合わせ先) (SPring-8に関すること) |
- 現在の記事
- スプリングエイトの高輝度X線が明らかにした昆虫飛翔筋の構造進化(プレスリリース)