細胞運動の“ブレーキ”の特性が明らかに(プレスリリース)
- 公開日
- 2010年07月07日
- BL26B1(理研 構造ゲノムI)
平成22年7月7日
国大学法人名古屋大学
独立行政法人理化学研究所
国立大学法人東北大学
独立行政法人科学技術振興機構
|
この度、名古屋大学大学院理学研究科の武田修一研究員、前田雄一郎教授を中心とする研究グループは、独立行政法人理化学研究所播磨研究所の似内靖先任研究員、名古屋大学大学院情報科学研究科の太田元規教授の研究グループ、及び東北大学大学院薬学研究科の山國徹准教授の研究グループと共同で、細胞運動を調節する重要なタンパク質である「アクチンキャッピングタンパク質」(CP)の活性制御の仕組みを明らかとすることに世界で初めて成功しました。 (論文) |
《背景》
私たちの身体を作っている細胞は、ある時期に活発に動きます。神経回路を形成するときには、神経細胞は“手”を伸ばし相手の神経細胞と“手”をつなごうとします。母胎内で身体が形成されるときには、個々の細胞は正しい配置を取るために、適切な場所へと移動します。免疫細胞は、細菌など異物が侵入した場所に駆けつけ、外敵を“飲み込み”ます。また制御不能な移動能を獲得したがん細胞は、別な組織に転移します。
これらすべての細胞運動は、たった一種類のタンパク質アクチンの離合集散によって引き起こされます(アクチンの分子運動)。細胞内ではアクチン分子は二つの状態をとります。すなわち、個々バラバラに存在する状態と、多くの分子が数珠のように互いに連なって(重合)アクチンフィラメントを作っている状態です(図1)。アクチンが重合し、細胞膜を内側から押すことによって、細胞は移動します。
細胞内には、アクチンの分子運動の速度を調節する多くの補助タンパク質が存在します。このような補助タンパク質の一つに、アクチンキャッピングタンパク質(CP)があります(図1)。CPはアクチンフィラメントの端に結合して、そこを塞ぐことによって重合を抑えます。CPが多すぎると、アクチンの重合が進まず、アクチンフィラメントができません。反対にCPが少なすぎると、細胞はアクチンフィラメントだらけになってしまいます。実際、細胞は時と場所に応じてCPの量を調節し、それによってアクチンフィラメントの量が調整され、それゆえ細胞運動の速度や強さを変動させることができます。つまりアクチンを細胞運動のエンジンに例えると、CPはブレーキに相当します。
さらに細胞は、このCPのブレーキ能力を調節する仕組みを備えています。本研究では、二種類のタンパク質、V-1(ブレーキの数を減らす)とCARMIL(ブレーキをアクチンフィラメントから外す)がどのようにCPを調節しているのかを、世界で初めて明らかにしました。
《研究成果のポイント》
1. アクチンキャッピングタンパク質(CP)とその作用を抑制するタンパク質V-1、及びCARMILそれぞれとの複合体の詳細な立体構造を解明した。
2. V-1はCP上のアクチンフィラメント結合部位を直接覆い隠すことによって、CPとアクチンフィラメントとの結合を不可能にする。その一方CARMILはV-1とは全く別の位置に結合する。
3. これまで“硬い”タンパク質であると考えられていたCPが、実は常にねじり運動を繰り返す“柔らかい”分子であることが判明した。
4. CARMILはCPを束縛し、ねじり運動を抑えることによって、CPをアクチンフィラメント端から解離させると結論された。
《研究の内容》
研究グループは今回の研究で大型放射光施設SPring-8※1の理研 構造ゲノム I ビームライン BL26B1を利用し、X線結晶構造解析法※2によりCPとV-1、及びCPとCARMILのそれぞれのタンパク質複合体の構造を解明することに成功しました。これによって、それぞれの分子同士がどこにどのように結合しているかを、詳しく知ることができました。
V-1はCP表面上の、アクチンフィラメント端と結合する部位を覆い隠すように結合していました(図2)。そのためV-1が結合したCPは、もはやアクチンフィラメント端には全く結合することができなくなる、つまりブレーキとしては働けなくなります。
一方CARMILはCP上のV-1とは反対側の位置、すなわちアクチンフィラメント端結合部位とは全く異なる場所に結合していました(図2)。これはCARMILがアクチンフィラメント端に結合しているCPにも結合できることを意味しています。
CARMILはCPをアクチンフィラメント端から解離させる(ブレーキを外す)ことが知られていますが、それでは、なぜ離れた場所に結合するにもかかわらず、CARMILはCPをアクチンフィラメントから外すことができるのでしょうか?この遠隔操作を実現するためには、CARMILが結合することによってCP全体の形に何らかの変化を起こす必要があります。今回の研究では、その変化の実態を解明することができました。
今回得られたいくつかのCPの構造を比較することによって、CPは大小二つの領域(ドメイン)から成り立っていることが示されました(図3)。二つの領域は互いにねじれるように揺れ動くことができるようになっており、それによってCPは分子の形を変えることができます。つまり、これまで“硬い”分子であると考えられていたCPが、実は“柔らかい”分子であることが判明しました。CPはアクチンフィラメント端にいったん接触すると、このねじり運動によって自身の形をアクチンフィラメント端にピッタリと合うように変化させるようです。CARMILはCPの二つの領域にまたがって結合しています。これらのことから、CARMILは二つの領域間のねじれ具合を変え、CPをアクチンフィラメント端とは合わない形に束縛するため、アクチンフィラメント端との結合を著しく弱めると考えられます(図4)。本研究では、この考え方を支持する証拠も得ることができました。
《成果の意義》
冒頭に述べたように、アクチンの分子運動はすべての細胞運動の駆動力であり、アクチンと共に働く補助タンパク質も、CPを含めてすべての細胞に共通であります。よって今回明らかとなったCPの作用を調節する仕組みは、細胞運動全般の仕組みを理解する上で非常に重要な知見となります。すなわち免疫、がん、神経発生といった重要な生命現象のメカニズムを理解する上での基礎となります。またV-1は心臓肥大の原因タンパク質の一つとして知られており、今回の結果が心臓病の治療薬の開発につながると期待されます。さらにCARMILがアクチンフィラメント端に結合したCPを外す仕組みは、CP自身の形の揺らぎを巧みに利用したもので、タンパク質相互作用の制御の方式一般を考える上で非常に興味深い例です。
《参考資料》
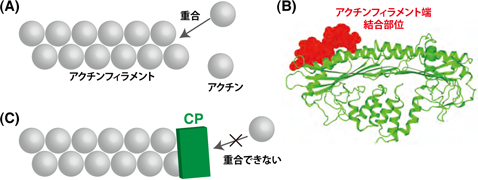
(A) 個々のアクチン分子が連なる(重合する)ことによって、アクチンフィラメントが形成される。
(B) アクチンキャッピングタンパク質(CP)の原子構造。楕円で囲った部分がアクチンフィラメント端結合部位。
(C) CPはアクチンフィラメント端を覆うことによって、重合を阻害する。
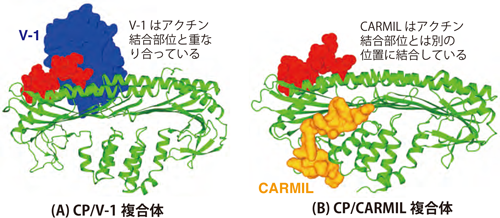
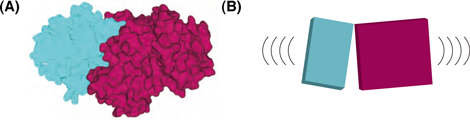
(A) CPは二つの領域(ドメイン;色分けして表示)から成り立っていることがわかった。
(B) 二つの領域は互いにねじれるように揺れ動く。この“柔らかさ” がアクチンフィラメント端に結合する時に重要であると考えられる。
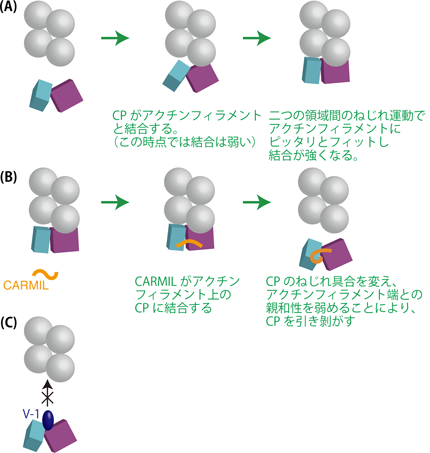
(A) CPは二つの領域間のねじれ運動によって、アクチンフィラメント端と強く結合する。
(B) CARMILはCPをアクチンフィラメント端とは合わない形に束縛することによって、CPを外す。
(C) V-1はCPのアクチン結合部位を直接覆い隠すので、V-1が結合したCPはアクチンフィラメントと結合できない。
《用語解説》
※1 大型放射光施設SPring-8(スプリングエイト)
SPring-8は兵庫県の播磨科学公園都市にある世界最高輝度の放射光を生み出す理研の施設。SPring-8の名前はSuper Photon ring-8GeV に由来。放射光とは、電子を光とほぼ等しい速度まで加速し、電磁石によって進行方向を曲げた時に発生する、細く強力な電磁波のこと。SPring-8では、この放射光を用いて、物理、化学、地学などの基礎研究から、ナノテクノロジー、バイオテクノロジーや産業利用まで幅広い研究が行われている。
※2 X線結晶構造解析法
タンパク質などの分子の構造を原子レベルで決める方法の一つ。まず調べたい分子を高純度に精製し、結晶を作成する。そこにX線を照射し、結晶中で規則的に並んだ分子にぶつかることによるX線の反射のパターンから、分子の形を計算する。一般的にタンパク質のような高分子は結晶になりにくいため、良い反射が得られる結晶化条件の探索には多大な労力を伴うことが多い。
(問い合わせ先) 名古屋大学大学院理学研究科 名古屋大学大学院情報科学研究科 東北大学大学院薬学研究科 理化学研究所播磨研究所 (報道に関する問い合わせ先) 独立行政法人理化学研究所 広報室 報道担当 (SPring-8に関すること) |

- 現在の記事
- 細胞運動の“ブレーキ”の特性が明らかに(プレスリリース)