昆虫類の口器の祖先型を解明 ~口器の進化に関する新しい考えを提唱~(プレスリリース)
- 公開日
- 2015年07月29日
- BL47XU(光電子分光・マイクロCT)
2015年7月29日
国立大学法人 筑波大学
公益財団法人 高輝度光科学研究センター
- 原始的な昆虫類である トビムシ目、コムシ目の口器をシンクロトロンµCTで詳細に検討した。
- 両目の口器は、共に同じ原理に基づく構造(口器の構造的連関、SMI)をもつことを明らかにした。
- 昆虫類の口器の祖先型は「SMI型」であることが判明したことで、SMIを持たない「噛み口」が原型であるとする従来の考えを否定し、昆虫類の口器の新たな進化像を提出した。
|
筑波大学生命環境系の町田龍一郎教授(菅平高原実験センター昆虫比較発生学研究室)および同研究室のアレクサンダー・ブランケ特別研究員、(公財)高輝度光科学研究センター利用研究促進部門の上杉健太朗副主幹研究員は、ドイツとスイスとの共同研究により、シンクロトロンµCTでの非破壊の機能形態学的研究を行い、昆虫類の口器の祖先型を明らかにし、昆虫類の口器の進化に関する新たな見方を提唱しました。 本研究は、日本学術振興会特別研究員奨励費および基盤研究C、筑波大学-ドイツ学術交流会(DAAD)パートナーシッププログラムの助成により、SPring-8、DESY、PSIの各放射光施設を利用して実施されました。 掲載論文 |
研究の背景
昆虫は地球上で最も多様化した生物であり、これまでに記載されている全動物種の約75%を占めています。昆虫がこのような多様化をとげた背景には「翅」や「変態」の獲得などによって多様な環境に進出できたことがあげられます。その背景として、多種多様な食物が利用可能な口器の多様化も大きな鍵を握っていました。したがって、口器の進化は昆虫類を理解する上できわめて興味深いテーマです。
昆虫の口器には、トンボやバッタなどに見られる普通の咀嚼型の口器である「噛み口」から、セミ、ハチの一部、チョウなどに見られる色々なタイプの「吸収口」、ハエなどの舐めるタイプの「舐め口」など、色々あります。口器は、上唇や、胸部の脚と由来を同じくする大顎、小顎、下唇(注3)などからなっています。昆虫に見られる口器の特殊化は、大顎、小顎、下唇などの顎構造が構造的に互いに連関することでもたらされました。時には顎構造が完全に結合することで、チョウに見られるようなストロー状の口を作ったりもします。このような顎の構造的な連携を「口器の構造的連関 Structural Mouthpart Interaction (SMI)」と呼びます。
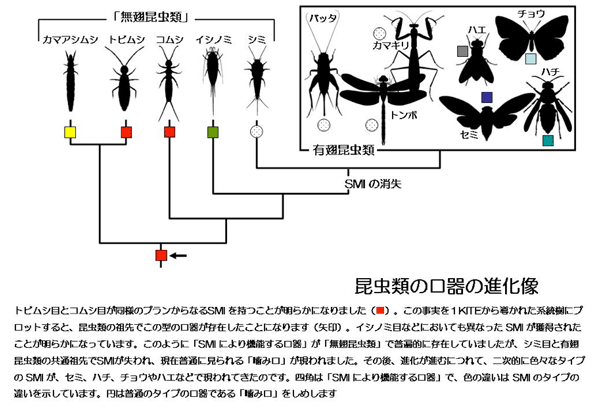
昆虫類は、まだ翅の獲得には至っていない、トビムシ目、カマアシムシ目、コムシ目、イシノミ目、シミ目が含まれる「無翅昆虫類」と総称されるグループと、昆虫類の大部分(約99%)を占める約25目からなる有翅昆虫類に分けられます。このSMIは、普通の「噛み口」であるトンボ目やバッタ目などの原始的な有翅昆虫類では存在せず、セミ目、チョウ目やハエ目など、より進化した有翅昆虫類のグループの特殊化した口器で知られています。そのため、昆虫類の祖先の口器は、もともとはSMIのない「噛み口」であり、収斂的にいろいろなグループでSMIが独立に獲得されたと理解されてきました。しかし、祖先的な昆虫類の研究がほとんど行われてこなかったことから、これまでの口器の進化に関する理解は、推測に基づく不十分なものでした。
研究内容と成果
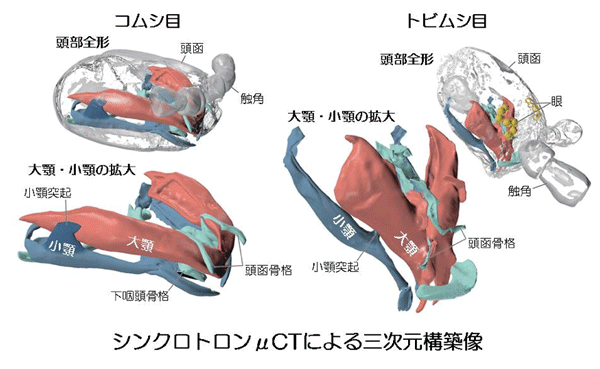
原始的な昆虫類である「無翅昆虫類」のトビムシ目(注4)、コムシ目(注5)の口器を、大型放射光施設SPring-8(注6)のBL47XU、ドイツ(DESY)およびスイス(PSI)の放射光施設においてシンクロトロンµCT(注7)法により詳細に観察し、両目の口器の詳細な三次元構築像を作成しました。これを基に、両目の口器がどのように機能するかに注目しつつ、構造を比較形態学的に検討しました。
解析の結果、以下のような事実が明らかとなり、昆虫の口器の新たな進化像が提出されました。
1)昆虫類の原始系統群であるトビムシ目、コムシ目の両目において、小顎突起(小顎の蝶咬節にある突起)の存在を確認。この突起により、小顎と大顎が連関することで口器が機能することから、両目の口器はSMIで機能することが明らかとなりました。
2)両目は、小顎突起による小顎と大顎の連関だけでなく、大顎が下咽頭骨格および頭函骨格により支持されて機能するという共通点を持つことも明らかになりました。すなわち、両目の口器はまったく相同(注8)のプランをもっていると結論づけられました。本論文の著者である筑波大学、ボン大学の研究者を含めた世界13カ国・43研究機関・101名の共同研究による、「昆虫トランスクリプトーム進化プロジェクト 1000 Insect Transcriptome Evolution (1KITE)」で構築された昆虫の系統樹(引用文献1)から、トビムシ目とコムシ目は単系統群(注9)を構成しないことが分かっています。したがって、この相同な口器は両目の共有派生形質(注10)とは理解できず、昆虫類の祖先で獲得された昆虫類の口器の祖先型(注11)(グラウンドプラン(注12))であると結論づけられました。
3)本論文の著者である筑波大学およびボン大学の研究者たちは、最近、同様に原始的系統群であるイシノミ目の口器にもSMIを見出しています(引用文献2)。したがって、昆虫類の口器のグラウンドプランは単純な「噛み口」であるとしてきた従来の理解は誤りであり、最初に生まれたのはSMIにより機能する口器であり、それからトンボやバッタなどに見られるような、SMIを失った「噛み口」が現われ、その後、二次的に多様なタイプのSMI、すなわち、セミ、ハチ、チョウなどの「吸収口」、ハエなどの「舐め口」が現れたことになります。
4)SMI による口器の多様化は、昆虫を多種多様な食物資源の獲得に導き、それが大きな要因の一つとなって昆虫の大繁栄がもたらされたと考えられます。SMI は進化した昆虫で後に獲得された新規形質であるという従来の理解は誤りであり、SMIは昆虫の祖先型ですでに存在していたことになります。昆虫類は出現の当初から、SMI の原理によって口器を様々に改変する「素養」を有していたことになり、実際、原始系統群においても、臨機応変に新たな「口器を開発」することで新たなニッチ(生態的地位)を開拓し、多様化を果たしていたものと考えられます。
今後の展開
複雑な成り立ちをしている構造は、それをもつ生物グループの進化を理解する上で有用です。昆虫類の口器はまさにこれに当たります。本研究による原始系統群の口器の詳細なシンクロトロンµCT解析により、従来の説とは大幅に異なる昆虫類の口器の進化像がもたらされました。カマアシムシ目など他の「無翅昆虫類」、進化上重要な位置にある有翅昆虫類の各グループの口器をターゲットに、本研究で採用したのと同様の非破壊のシンクロトロンµCTを用いた詳細な検討がなされることになるでしょう。そして、このような研究成果と地球科学分野からの地質時代の環境推定が統合されれば、昆虫類の進化に関するより詳細な理解が得られることになります。
参考図

《用語解説》
注1)口器:
節足動物、昆虫類の口は、上唇、大顎、小顎、下唇、下咽頭などさまざまな構造から構築されています。このような多様な構造により形作られている昆虫類などの口を口器と呼びます。
注2)口器の構造的連関 (Structural Mouthpart Interaction、SMI):
普通の昆虫が持っている「噛み口」の場合、口器の主要素である大顎、小顎などは互いに構造的に連関することなく別々に動くことで機能します。これに対して、大顎や小顎などが構造的につながることで機能する場合があります。各要素が構造的に連関することで機能する口器における、口器の連携をこのように呼びます。
注3)大顎、小顎、下唇:
昆虫類などの節足動物の体は、体節が連続した構造が基本で、個々の体節はそれぞれが1対の脚(付属肢)をもっています。昆虫の脚は胸部にあり、胸部は3体節で構成されていることから、3対の歩脚をもっています。腹部の脚は一般に退化しています。頭部は複数の体節が合体しており、それぞれの体節の脚は触角や摂食に用いられる大顎、小顎、下唇などにそれぞれ特殊化しています。
注4)トビムシ目:
土壌に多数生息する1~数ミリの「無翅昆虫類」の一群。落葉、カビなどを食べ、マルトビムシ亜目とフシトビムシ亜目の2亜目があります。写真はフシトビムシ亜目のデガトゲトビムシ。
注5)コムシ目:
土壌に生息する数ミリ程度の「無翅昆虫類」の一群。2亜目からなり、ナガコムシ亜目は落葉やカビ、ハサミコムシ亜目は小さな土壌動物を餌とします。写真はハサミコムシ亜目のヤマトハサミコムシ。
注6)大型放射光施設 SPring-8:
兵庫県の播磨科学公園都市にある世界最高の放射光を生み出す国立研究開発法人理化学研究所の施設で、その管理運営は公益財団法人高輝度光科学研究センターが行っています。SPring-8 の名前は Super Photon ring-8 GeV
に由来。放射光とは、電子を光とほぼ等しい速度まで加速し、電磁石によって進行方向を曲げた時に発生する、細く強力な電磁波のこと。
注7)シンクロトロンµCT:
大型円形加速器シンクロトロンから得られる高輝度放射X線を照射、サンプルの高分解能の断層像を獲得し、コンピューターで高解像度の3次元構築を行なう技術です。分解能が高いだけでなく、解剖などと違い非破壊で解析できるのも大きなメリットです。構造同士の位置関係、連関などの情報が保たれるのです。
注8)相同:
構造などの類似が共通の祖先の由来に基づくものである場合、その類似を相同といいます。
注9)単系統群:
単一の祖先に由来するすべての種群を含んだ系統群。参考図にある系統樹において、トビムシ目とコムシ目の共通祖先(この系統樹では、最基部の幹にあたります)に由来する種群は両目以外のすべての昆虫類も含むので、両目は単系統群とはいえません。
注10)共有派生形質:
単系統群である系統群が分岐したときの共通祖先が持っていた派生形質のこと。
注11)祖先型:
祖先と想定される生物群のこと、あるいはそれが持っていると想定される特徴群。
注12)グラウンドプラン:
系統群の本質的な特徴。
引用文献
1 Misof, B. et al. (2014) Phylogenomics resolves the timing and pattern of insect evolution. Science, 346(6210): 763-767.
2 Blanke, A., R. Machida, N. U. Szucsich, F. Wilde and B. Misof (2014) Mandibles with two joints evolved much earlier in the history of insects: Dicondyly is a synapomorphy of bristletails, silverfish and winged insects. Systematic Entomology, 40(2): 357-364.
|
《問い合わせ先》 上杉 健太朗(うえすぎ けんたろう) (SPring-8に関すること) |
- 現在の記事
- 昆虫類の口器の祖先型を解明 ~口器の進化に関する新しい考えを提唱~(プレスリリース)