自然免疫における核酸センサーTLR8のリガンド認識および活性化機構の解明 -抗ウイルス薬、がん免疫賦活剤の開発に道-(プレスリリース)
- 公開日
- 2013年03月22日
- BL41XU(構造生物学I)
- BL45XU(構造生物学 III)
2013年3月22日
東京大学
発表のポイント
• どのような成果を出したのか
ウイルス由来のRNAを感知して炎症、抗ウイルス応答を引き起こすToll Like Receptor 8 (TLR8)受容体の、リガンド非結合型(不活性化状態)とリガンド結合型(活性化状態)の詳細な三次元構造を明らかにしました。
• 新規性(何が新しいのか)
TLR8は、リガンドを認識することでその2量体構造が再構成され細胞内にシグナルを伝えることを明らかにしました。
• 社会的意義/将来の展望
重要な創薬ターゲットとなるTLR8の詳細な三次元構造が分かったことで、TLR8の活性を制御する抗ウイルス薬やワクチンなどの開発につながることが期待されます。
|
東京大学大学院薬学系研究科の清水敏之 教授、大戸梅治 助教、同大薬学部学生 丹治裕美および東京大学医科学研究所の三宅健介 教授、柴田琢磨 特任助教の研究グループは、ウイルスの侵入を感知して免疫系を活性化するTLR8受容体の詳細な三次元構造を世界で初めて解明しました。 (論文) |
研究の背景と経緯
細菌やウイルスなどの病原体に対する防御機構として、我々の体には自然免疫機構が備わっています。TLRは、自然免疫機構において重要な役割を果たすI型膜タンパク質受容体です。細菌やウイルスなどが持っている特有の分子パターンは、異物としてTLRによって感知され、我々の体に炎症応答や抗ウイルス応答を引き起こします。これまでにヒトでは10種類のTLRが報告されており、それぞれが異なる分子パターンを見分けています。TLRは細胞外領域で分子パターンを認識して、細胞内領域を通じて、自然免疫応答を引き起こしています。
TLR8(およびTLR7)は、A型インフルエンザウイルスなどに由来する一本鎖RNAを感知して、炎症性サイトカイン、I型インターフェロンの産生を促して炎症、抗ウイルス応答を引き起こします。一方で、自身のRNAにも反応して異常な炎症応答を引き起こし得ることから、自己免疫疾患などの病態と関係する可能性も考えらます。TLR7/8は、合成低分子化合物によっても活性化することが知られており、これらの化合物は、免疫賦活剤として臨床に応用されています。これまで、TLR7/8がどのようにしてRNAまたはこれらの低分子化合物により活性化され細胞内へ情報を伝えるのかについての具体的な機構は不明でした。
研究の内容
本研究グループは、TLR8によるリガンドの認識とシグナル伝達機構をX線結晶解析の手法を用いて明らかにするために、ヒトTLR8の細胞外領域の大量調製に成功しました。得られたタンパク質を用いて結晶化を行い、最終的に、リガンドが非結合型TLR8と3種類の低分子リガンド(CL097、CL075、R848)とTLR8との複合体の結晶構造を明らかにすることに成功しました。結晶からの回折像の測定には、大型放射光施設SPring-8および高エネルギー加速器研究機構Photon Factoryの強力なX線を使用し、リガンド非結合型を2.3 Å、結合型を2.0~2.7 Åという高い分解能での構造決定に成功しました。
構造解析の結果、リガンド結合型と非結合型は、いずれも2分子のTLR8のC末端部分が中央に配置したm字型となるように集まった2量体を形成していることが分かりました(図1)。リガンドは2回対称軸で関係づけられる2ヶ所に結合しており、TLR8との間に特徴的な相互作用が形成されていることが分かりました(図2)。リガンド非結合型と結合型の2量体間では2量体を形成する界面が異なり、後者では2つのプロトマーがより近接していました。それに伴い、2量体を形成するプロトマーのC末端同士の距離は、53 Å(リガンド非結合型)から~30 Å(結合型)と接近していました。リガンドの結合によりTLR8の2量体が再構成され、TLRドメインのC末端及び細胞内ドメインが接近することでシグナルが下流へ伝えられると考えられます(図3)。これまで、TLRの活性化機構として、単量体TLRがリガンドとの結合に伴い2量体化することで活性化するという機構が考えられていましたが、今回の結果はTLRの新たな活性化機構を実証するものといえます。
今後の展開
TLRの中でも、病原体由来のDNAやRNAなどの核酸を認識するTLR7、TLR8、TLR9は、抗ウイルス薬やワクチンのアジュバントなどの創薬上重要なターゲットとして注目を集めています。特にTLR7やTLR8は、TLRのリガンドの中でも最も小さい分子量200~300程度の化合物で活性化されます。これは創薬上非常に大きなメリットだと考えられます。しかしながら、これらTLRは、TLRの中でも最もアミノ酸配列が長く、また、高度に糖鎖修飾されていることもあって、結晶化可能な量のタンパク質を調製することが困難で、これまでタンパク質レベルでの具体的な活性化機構は不明でした。今回、その活性化機構が三次元構造レベルで明らかになったことから、TLR8をターゲットとした化合物をコンピュータ上で設計および評価することが可能となります。
本研究は、科学研究費補助金、公益財団法人武田科学振興財団助成金(薬学奨励研究)、公益財団法人持田記念医学薬学振興財団助成金などの外部資金支援を受けて行われたものです。
《参考図》
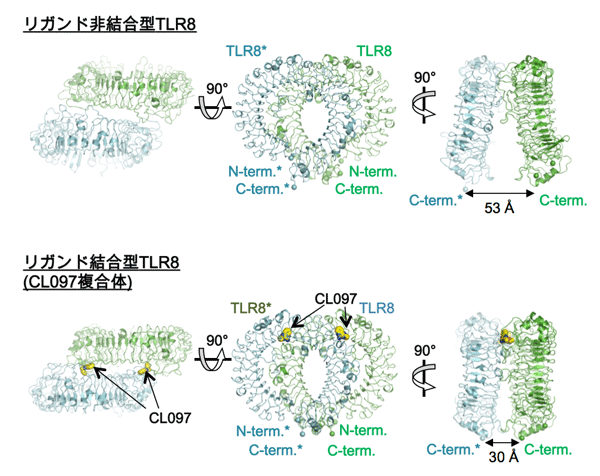
(上図)リガンド非結合型のTLR8。(下図)リガンド結合型のTLR8 (CL097複合体)。2量体を形成するTLR8分子の一方を緑色で、他方を青色で示している。リガンドは2量体に2ヶ所で結合している。リガンド結合型では、C末端同士の距離が近くなっている。
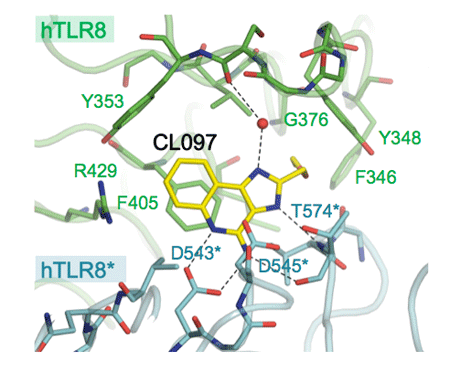
CL097複合体について示している。リガンド(黄色)は、緑と青で示す2分子のTLR8の間に結合している。水素結合を点線で示している。CL075複合体、R848複合体もほぼ同様の格好で結合していた。
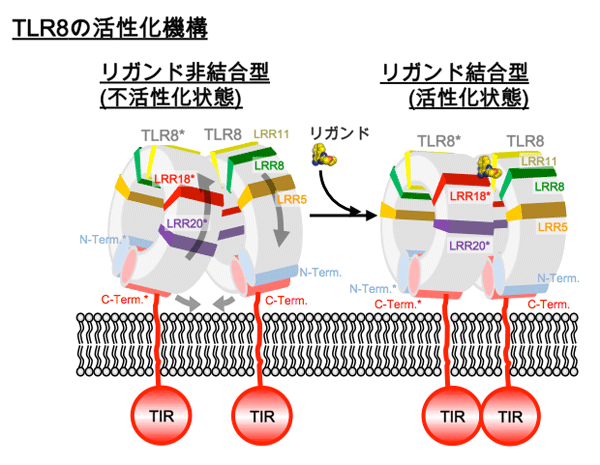
TLR8の活性化のモデル図。リガンド結合によりTLR8のC末端側が近接することで細胞内TIRドメインが近接し、細胞内へと情報が伝わるものと考えられる。
|
《問い合わせ先》
(SPring-8に関すること) |
- 現在の記事
- 自然免疫における核酸センサーTLR8のリガンド認識および活性化機構の解明 -抗ウイルス薬、がん免疫賦活剤の開発に道-(プレスリリース)