脱窒過程における銅含有亜硝酸還元酵素と電子供与体蛋白質の過渡的電子伝達複合体の立体構造を世界で初めて解明-有害窒素酸化物を削減する戦略の確立に役立つと期待-(プレスリリース)
- 公開日
- 2009年11月05日
- BL44XU(生体超分子複合体構造解析)
2009年11月5日
大阪大学
|
大阪大学大学院理学研究科の鈴木晋一郎教授・野尻正樹助教の研究グループは、大型放射光施設SPring-8の大阪大学蛋白質研究所専用ビームラインの生体超分子構造解析ビームラインBL44XUを用いて、世界で初めて銅含有亜硝酸還元酵素とその電子供与体蛋白質との過渡的電子伝達複合体の立体構造を解明することに成功しました。なお、本研究の成果は、11月5日発行の英国雑誌「Nature」に掲載されます。 (論文) |
研究の背景及び成果
蛋白質分子間の電子伝達反応は、「光合成系」、「呼吸鎖」、「各種代謝系」など生体内の多くの反応系に存在するプロセスであり、生物の生存に欠くことのできない反応です。地球上の窒素循環に深く関わる「脱窒」(NO3-→NO2-→NO→N2O→N2)を行う微生物においても同様に、硝酸イオンから続く段階的還元反応を触媒する各種還元酵素はその触媒反応に必要な電子を蛋白質やキノン分子等から分子間電子伝達反応により効率よく得ています。一般に、電子伝達パートナー分子同士によって形成される過渡的な複合体は弱く短寿命な一過性のものであり、それは生体中において分子間電子伝達反応を効率よく繰り返し行うために必要な物理化学的特性です。しかしながら、その不安定さからこれまでに複合体形成時の立体構造を高分解能で解析した例は非常に少なく(プロテイン・データ・バンク(PDB)にも十数例しかない)、その反応メカニズムを詳細に理解することは困難とされています。
大阪大学大学院理学研究科の鈴木晋一郎教授・野尻正樹助教らは、脱窒過程の中で特に温室効果ガスの1つとして注目される亜酸化窒素(N2O)の前駆物質である一酸化窒素(NO)を触媒反応生成物とする亜硝酸還元酵素※1とその電子供与体蛋白質であるチトクロムc-551間で起こる蛋白質分子間電子伝達反応に着目し、その複合体の構造を大型放射光施設SPring-8の生体超分子複合体構造解析ビームラインBL44XUを利用したX線結晶構造解析※2によって1.7Åの分解能で決定しました。
構造決定された複合体は分子量約37-kDaのサブユニット3つから成るホモ三量体構造の亜硝酸還元酵素分子の上に1つのチトクロムc-551分子が結合していました(図1)。電子の授受が起こるチトクロムc-551のヘムc※3と亜硝酸還元酵素のタイプ1銅※4部位は10.5Åの距離に位置し、お互いの酸化還元反応中心が最も近づくような配向で接していました。両分子間で形成された相互作用界面では、通常、分子表面に存在する溶媒水分子が両分子間の相互作用により排除された状態になっており、その界面中心には疎水的な環境が過渡的に形成される事がわかりました(図2)。相互作用界面のアミノ酸残基とその位置、そして水分子の位置を特定できたことで、蛋白質分子間電子伝達反応に関する研究・考察が飛躍的に進むものと期待されます。
2008年にオランダ・ライデン大学の研究グループが他の菌株由来の銅含有亜硝酸還元酵素とその電子供与体蛋白質シュードアズリンの過渡的相互作用についてNMR※5を用いて解析しています。その解析から示唆される複合体構造と今回決定した複合体構造を比較すると、それぞれの電子供与体がチトクロムc-551とシュードアズリンで全く異なる構造をした蛋白質であるにも関わらず、亜硝酸還元酵素のほぼ同じ部位に相互作用していることがわかりました。これは、蛋白質分子間電子伝達反応における擬特異的分子認識メカニズムといわれるものであり、今回の立体構造決定により亜硝酸還元酵素の電子伝達パートナー分子認識機構における擬特異性の構造的基盤を明らかにすることができました。また、今回の立体構造を基にした部位特異的変異体を用いた解析から、電子移動反応過程における鍵となるアミノ酸の1つがタイプ1銅近傍のメチオニン残基であることも突き止めました(図3)。
本研究における亜硝酸還元酵素とチトクロムc-551の電子伝達系は、地球上の窒素循環のバランスを保つために重要な役割を担う「脱窒」プロセスの一端をなします。本研究により、これまで解析が困難であった両蛋白質分子間の分子認識メカニズムと分子間電子伝達メカニズムをより正確に理解する事ができ、原子レベルで地球上の窒素レベルをコントロールするデバイス開発への可能性を広げることができます。また、近年の地球温暖化への危惧から、二酸化炭素と肩を並べる温室効果ガスとして脱窒プロセスから発生する「亜酸化窒素(N2O)ガス」が注目されており、地球上の窒素循環をコントロールする技術は二酸化炭素に続き期待される次世代技術と言っても過言ではありません。そして、蛋白質ー蛋白質の分子間相互作用は、地球上の生命(細胞)全てが持つ機能であり、その強さ、特異性、結合/解離バランスがその分子環境に応じて巧みにコントロールされています。この機能を発揮する1つの要因は“蛋白質ならでは”の「柔軟性」であり、本研究成果は、そういった柔軟性を取り入れた分子スイッチ等への開発技術へも大きく貢献し得ると期待されます。
<参考資料>
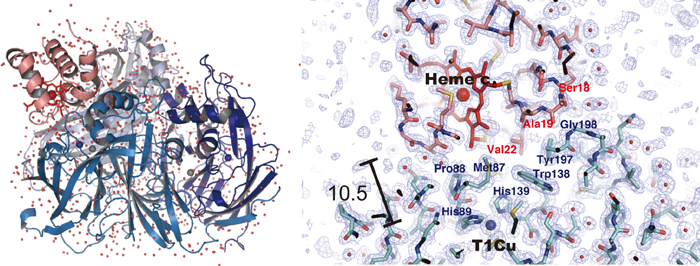 図1 銅含有亜硝酸還元酵素とチトクロムc-551複合体の全体構造(左)と相互作用部位(右)
図1 銅含有亜硝酸還元酵素とチトクロムc-551複合体の全体構造(左)と相互作用部位(右)亜硝酸還元酵素分子(ホモ三量体)の上に1分子のチトクロムc-551(ピンク色リボン描画)がのっている。チトクロムc-551のヘムcと亜硝酸還元酵素のタイプ1銅部位(図内Heme cとT1Cu)が10.5Åの距離まで近づいている。
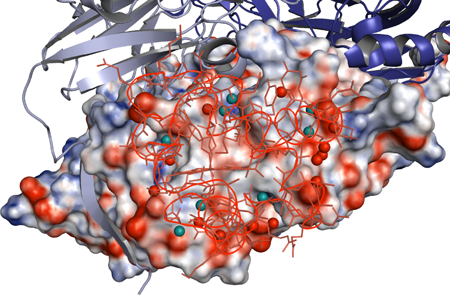 図2 相互作用界面の水分子の位置
図2 相互作用界面の水分子の位置相互作用界面の水分子を球(水色と赤色)で描画している。水色は両分子間で水素結合を形成している水分子を表す。赤色表示の水分子はどちらか一方の分子と水素結合している。相互作用界面の中心には水は存在しない。チトクロムc-551が相互作用している亜硝酸還元酵素サブユニットの分子表面における電荷分布も表示している(赤:負、青:正)。チトクロムc-551分子をワイヤー描画で表示している。
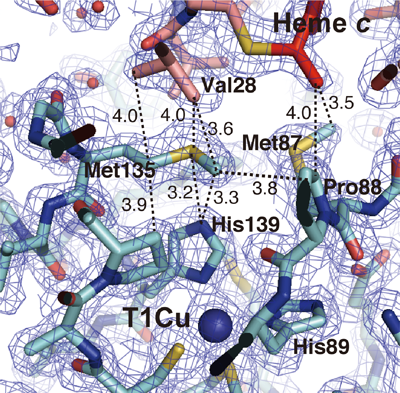 図3 分子間電子伝達反応で鍵となるメチオニン残基
図3 分子間電子伝達反応で鍵となるメチオニン残基亜硝酸還元酵素のメチオニン135番はチトクロムc-551が結合すると両分子の酸化還元中心(c型ヘムとタイプ1銅)により挟まれた位置で固定された構造をとる。このメチオニンをセリンに置換すると両分子間の電子移動速度定数が約10分の1に低下する。
<用語解説>
※1 亜硝酸還元酵素
亜硝酸塩(イオン)の還元を触媒する酵素で異化型と同化型がある。異化型酵素は脱窒菌に存在し、補欠分子にヘムを持つタイプと銅を持つタイプがある。
※2 X線結晶構造解析
物質を3次元的に規則正しく並べる事で結晶を作り、その結晶にX線を照射し、回折パターンとその強度から物質の構造を解析する方法。蛋白質の構造決定では多く用いられている。
※3 ヘムc
チトクロムc等の補欠分子族でポルフィリンcの鉄錯体。3,8位のα-システイニルチオエーテル結合で蛋白質骨格に共有結合している。
※4 タイプ1銅
一般に2つのヒスチジンと1つのシステイン、そして1つのメチオニン残基で蛋白質内部に保持された単核銅結合部位のこと。まれに、さらに蛋白質主鎖のカルボニル酸素が配位したもの、メチオニンの代わりにグルタミンが配位したもの、そしてメチオニンのないものが存在する。システインの硫黄原子から銅原子へのLigand-to-Metal Charge Transfer (LMCT)により強い青色を呈することから「ブルー銅」とも呼ばれる。
※5 NMR
分子や原子を調べる方法の1つである核磁気共鳴(Nuclear Magnetic Resonance)のこと。タンパク質の立体構造の2割程度がこの装置を用いて決定されている。溶液で構造決定できるので動的構造、また比較的弱い分子結合の情報が得られる。
|
(問い合わせ先) (SPring-8に関すること) |
- 現在の記事
- 脱窒過程における銅含有亜硝酸還元酵素と電子供与体蛋白質の過渡的電子伝達複合体の立体構造を世界で初めて解明-有害窒素酸化物を削減する戦略の確立に役立つと期待-(プレスリリース)